

1.유전자 설명
KDM2b는 F-box 단백질 계열의 구성 요소를 코드화하고, F-box 단백질 계열은 약 40개 아미노산의 특징적인 시퀀스를 가지고 있습니다. F-box 단백질은 유비퀴틴 단백질 연결 효소 복합체 SCF(SKP1-cullin-F-box)의 네 가지 하위 단위 중 하나를 구성하며 인산화 매개 유비퀴틴화 기능을 가지고 있습니다. F-box 단백질: WD-40 도메인을 포함하는 Fbws 유형, 류신 반복 구조를 포함하는 Fbls 유형, 그리고 마지막 유형인 Fbxs는 서로 다른 단백질과 상호 작용하는 시퀀스를 포함하고 있지 않거나 일부 식별되지 않은 시퀀스를 포함하는 세 가지 유형으로 나뉩니다. Kdm2b에 의해 코드화된 단백질은 Fbls 유형에 속합니다. Kdm2b에는 여러 개의 대체 스플라이싱 전사체가 있는 것으로 밝혀졌으며, 이데 따라 일부 전사체의 기능은 아직 완전히 결정되지 않았습니다[1].
2.질병 설명
KDM2b와 관련된 질병에는 코헨증후군 및 유아 경련이 있습니다. 이 단백질과 관련된 기능 경로에는 염색질의 3차원 구조 조절 및 염색질의 후성적 변형이 포함됩니다. 이 유전자와 관련된 GO 주석의 기능에는 rRNA 결합 능력과 히스톤 데메틸라제 활성이 포함됩니다. 이 유전자의 중요한 상동 유전자는 KDM2A입니다[2].
3.마우스 모델 표형 설명
유전자가 결실된 이 대립형질 동형접합 마우스(긴 전사체)는 출생 후 뇌 노출, 배아 또는 출생 후 치사율, 비정상적인 안구 조직, 꼬리 말림, 정자 과소증, 세포자멸사 증가, 신경세포 전구체 증식 증가를 보였습니다[3].
이전 연구에 따르면 FBXL10(KDM2b)은 다음 두 가지 주요 전사체를 발현하는 것으로 나타났습니다. FBXL10-1은 더 긴 전사체로, 이 전사체에서 번역된 단백질은 N-말단에 히스톤 데메틸라제 도메인이 있고, C-말단에 F-box, CXXC, PHD, RING 및 류신 반복 도메인이 있습니다. FBXL10-2는 더 짧은 전사체이고 전사는 다른 엑손에서 시작되며 이 전사체 번역된 단백질에는 히스톤 데메틸라제 도메인이 없지만 다른 모든 도메인은 유지합니다.
연구자들은 또한 FBXL10-1과 FBXL10-2 결실 유형이 현저하게 다른 표현형을 가지고 있음을 발견했습니다. FBXL10-2 결실 마우스는 전뇌 노출, 비정상적인 안구 조직 또는 FBXL10-1 돌연변이체와 같은 꼬리 말림을 보이지 않습니다. 그와 대조적으로, FBXL10-2 생존 개체는 저신장 및 두개안면 발육 이형성증의 특성을 갖는 반면, 출생 전에 사망한 개체는 두개안면 및 신경관 발달 이상 현상이 나타났습니다(그림1). FVB와의 역교배 후, FBXL10∆−2/∆−2를 가진 태아도 출생 시 척추 지지 균열과 눈꺼풀 이상 현상을 보였습니다(그림1)[4].
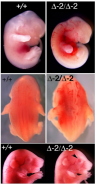
그림1. E12.5 배아(제 1 라인)와 E17.5 배아(제 3 라인)에서 FBXL10-2 대립유전자가 동형 결손된 상태에서 두개안면 이상을 보였습니다. 유전자 결실 마우스 태아의 두부 전면이 압력을 받는 현상이 뚜렷했고 출생 시에 눈을 뜨고 있는 표현형(하단에 검은색 화살표)도 있었습니다. Δ-2/Δ-2 배아(중간 라인)에서 굴곡된 신경관을 관찰할 수 있습니다.[4]
연구원들은 FBXL10의 두 하위 유형을 삭제함으로써 FBXL10T 동형 접합 모델을 편집하고, 이 모델을 사용하여 다른 성별 간의 표현형을 연구했습니다. 야생형과 비교하면, 이 배아는 그림2와 같이 더욱 심각한 표현형과 더욱 분명한 성별 간의 차이를 보였습니다. 배아는 10.5일째에 발달을 멈추고 Fbxl10∆−2/∆-2에서 발견되는 신경관 꼬임을 포함한 여러 가지 심각한 발달 이상이 발생했습니다.그림2는 FBXL10-1과 FBXL10-2 유전자의 결실에 의해 매개되는 성별 차이를 나타낸 것으로, 발달이 가장 낮은 암컷 배아와 발달이 가장 높은 수컷 배아가 거의 같은 단계에서 발육을 멈추는 것을 관찰할 수 있습니다[4].
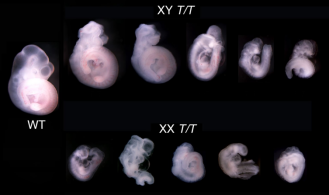
그림2. Fbxl10T/T 배아의 성별 차이 표현형의 해부학적 도표. 배아기 10.5일째에 배아를 채취하여 Sry 유전자로 성별을 확인했습니다.[4]
Fbxl10 결실 ES 세포는 계속 줄기를 유지함
연구자들은 배반포에서 Fbxl10T 대립유전자에 대해 동형접합성인 ES 세포를 분리하고 배양 배지에서 확장시켰습니다. 연구자들은 돌연변이 ES가 다능성 표기물 NANOG 및 OCT4를 발현할 수 있고(그림3a), 야생형 배아에서 키메라가 될 수 있고 모든 조직의 성장과 분화를 지원할 수 있다는 것을 발견했습니다(그림3b). DNA 메틸트랜스퍼라제 유전자 발현의 분석은 Fbxl10 T/T ES 세포가 미분화 ES 세포의 특성을 가지고 있음을 추가로 증명했으며, Fbxl10T/T 세포는 배우자 형성의 특정 단계에서 미분화 ES 세포 및 생식 세포에 고유한 Dnmt3a(Dnmt3A2) 및 Dnmt3L을 발현합니다. 그 외에도, 야생형 ES 및 Fbxl10T/T ES에서 Dnmt1 및 Dnmt3B의 발현 수준도 매우 유사합니다.
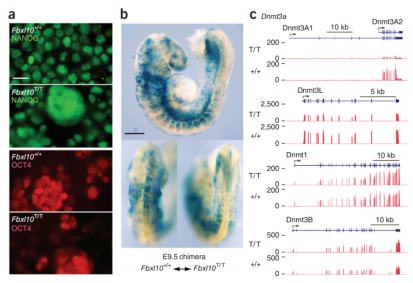
그림3. Fbxl10 T/T 배아줄기세포는 다능성과 배아줄기세포의 특성을 유지하고 있습니다. (A) 다능성 표기물 NANOG(상단) 및 OCT4(하단)의 발현. (B) WT 배반포에 주입한 후 돌연변이 ES 세포는 E9.5 배아의 모든 조직 발달을 지원하여 돌연변이 ES 세포의 다능성을 나타냅니다. (C) DNA 메틸트랜스퍼라제 발현 프로파일 분석. ES 세포 특이적 Dnmt3A2 및 Dnmt3L은 야생형 및 Fbxl10T/T ES 세포에서 발현되고, Dnmt3L은 분화된 세포에서 발현되지 않습니다. 그 외에도, 돌연변이체 및 야생형 ES 세포에서 Dnmt1 및 Dnmt3B의 발현 프로파일은 일관적입니다[5].
Fbxl10 유전자 이상은 전뇌 기형을 유발할 수 있음
Fbxl10+/- 이형접합 배아는 폐쇄되지 않은 중뇌 및 후뇌 신경관뿐만 아니라 전뇌 기형을 보였습니다. 비정상 전뇌를 가진 모든 Fbxl10-/- 마우스는 출생 직후 중추 신경계 장애로 사망으며, 또한 E18.5에서 뇌출혈 및 체형 감소를 보였습니다(그림4G 및 H). 그 밖에도, Fbxl10-/-의 배아에서도 안구 조직 결손과 꼬리 말림 특징을 나타내었으며(그림4F), 비정상 비율은 각각 40% 및 10%였습니다[6].
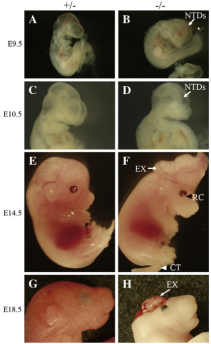
그림4. Fbxl10 결실 배아 표현형. (A–H) Fbxl10-/- 및 대조군의 다른 단계에서 배아의 측면도. A-D: 9.5일 및 10.5일의 배아 측면도로, 대조군 배아(A 및 C)의 신경관은 닫혀 있지만 돌연변이 배아의 신경 주름은 여전히 닫히지 않았습니다(B 및 D, 화살표). E 및 F: 배아 단계의 14.5일에서, 중뇌 이형성증(E)(화살표), 안구 조직 결손(검은색 삼각형) 및 꼬리 말림(흰색 삼각형)를 포함한 일부 Fbxl10-/- 배아(F) 표현형입니다. G 및 H: 배아기 18.5일째에서 Fbxl10-/- 배아(H)는 대조군 배아(G)보다 작으며 전뇌출혈로 전신이 창백합니다.[6]
4.결론
KDM2b는 염색질의 구조와 히스톤의 후성 유전학적 변형을 조절할 수 있는 중요한 단백질로 간주되어, 연구자들이 점점 더 깊이 연구하고 있습니다. Kdm2b는 신경과 뇌의 발달에 중요한 역할을 하며, 향후 연구에서는 해당 단백질이 분자 수준에서 모델을 조절하는 방식과 염색질 조직 및 히스톤 변형에 이 단백질이 미치는 영향에 초점을 맞출 수 있습니다.



영업일 기준 1-2일 내에 답변해 드리겠습니다.



